Plant hormone

Lack of the plant hormone auxin can cause abnormal growth (right)
Plant hormones (also known as phytohormones) are signal molecules produced within plants, that occur in extremely low concentrations. Plant hormones control all aspects of growth and development, from embryogenesis, the regulation of organ size, pathogen defense, stress tolerance and through to reproductive development. Unlike in animals (in which hormone production is restricted to specialized glands) each plant cell is capable of producing hormones.[1][2][3] The term 'phytohormone' was coined by Went and Thimann and used in the title of their book in 1937.[4]
Phytohormones are found across the plant kingdom, and even in algae, where they have similar functions to those seen in higher plants.[5] Some phytohormones also occur in microorganisms, such as unicellular fungi and bacteria, however in these cases they do not play a hormonal role and can better be regarded as secondary metabolites.[6]
Contents
1 Characteristics
2 Classes
2.1 Abscisic acid
2.2 Auxins
2.3 Brassinosteroids
2.4 Cytokinins
2.5 Ethylene
2.6 Gibberellins
2.7 Jasmonates
2.8 Salicylic acid
2.9 Strigolactones
2.10 Other known hormones
3 Potential medical applications
4 Hormones and plant propagation
4.1 Seed dormancy
5 References
6 External links
Characteristics

Phyllody on a purple coneflower (Echinacea purpurea), a plant development abnormality where leaf-like structures replace flower organs. It can be caused by hormonal imbalance, among other reasons.
The word hormone is derived from Greek, meaning set in motion. Plant hormones affect gene expression and transcription levels, cellular division, and growth. They are naturally produced within plants, though very similar chemicals are produced by fungi and bacteria that can also affect plant growth.[7] A large number of related chemical compounds are synthesized by humans. They are used to regulate the growth of cultivated plants, weeds, and in vitro-grown plants and plant cells; these manmade compounds are called plant growth regulators or PGRs for short. Early in the study of plant hormones, "phytohormone" was the commonly used term, but its use is less widely applied now.
Plant hormones are not nutrients, but chemicals that in small amounts promote and influence the growth,[8] development, and differentiation of cells and tissues. The biosynthesis of plant hormones within plant tissues is often diffuse and not always localized. Plants lack glands to produce and store hormones, because, unlike animals—which have two circulatory systems (lymphatic and cardiovascular) powered by a heart that moves fluids around the body—plants use more passive means to move chemicals around their bodies. Plants utilize simple chemicals as hormones, which move more easily through their tissues. They are often produced and used on a local basis within the plant body. Plant cells produce hormones that affect even different regions of the cell producing the hormone.
Hormones are transported within the plant by utilizing four types of movements. For localized movement, cytoplasmic streaming within cells and slow diffusion of ions and molecules between cells are utilized. Vascular tissues are used to move hormones from one part of the plant to another; these include sieve tubes or phloem that move sugars from the leaves to the roots and flowers, and xylem that moves water and mineral solutes from the roots to the foliage.
Not all plant cells respond to hormones, but those cells that do are programmed to respond at specific points in their growth cycle. The greatest effects occur at specific stages during the cell's life, with diminished effects occurring before or after this period. Plants need hormones at very specific times during plant growth and at specific locations. They also need to disengage the effects that hormones have when they are no longer needed. The production of hormones occurs very often at sites of active growth within the meristems, before cells have fully differentiated. After production, they are sometimes moved to other parts of the plant, where they cause an immediate effect; or they can be stored in cells to be released later. Plants use different pathways to regulate internal hormone quantities and moderate their effects; they can regulate the amount of chemicals used to biosynthesize hormones. They can store them in cells, inactivate them, or cannibalise already-formed hormones by conjugating them with carbohydrates, amino acids, or peptides. Plants can also break down hormones chemically, effectively destroying them. Plant hormones frequently regulate the concentrations of other plant hormones.[9] Plants also move hormones around the plant diluting their concentrations.
The concentration of hormones required for plant responses are very low (10−6 to 10−5mol/L). Because of these low concentrations, it has been very difficult to study plant hormones, and only since the late 1970s have scientists been able to start piecing together their effects and relationships to plant physiology.[10] Much of the early work on plant hormones involved studying plants that were genetically deficient in one or involved the use of tissue-cultured plants grown in vitro that were subjected to differing ratios of hormones, and the resultant growth compared. The earliest scientific observation and study dates to the 1880s; the determination and observation of plant hormones and their identification was spread-out over the next 70 years.
Classes
Different hormones can be sorted into different classes, depending on their chemical structures. Within each class of hormone the exact structures vary, but they have similar physiological effects. Initial research into plant hormones identified five major classes: abscisic acid, auxin, cytokinins, ethylene and gibberellins.[11] This list was later expanded and brassinosteroids, jasmonates, salicylic acid and strigolactones are now considered as major plant hormones. Additionally there are also several other compounds that fulfill a similar function to the major hormones, but their status as bone fide hormones is still debated.
Abscisic acid
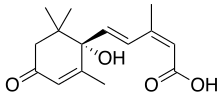
Abscisic acid
Abscisic acid (also called ABA) is one of the most important plant growth regulators. It was discovered and researched under two different names before its chemical properties were fully known, it was called dormin and abscicin II. Once it was determined that the two compounds are the same, it was named abscisic acid. The name "abscisic acid" was given because it was found in high concentrations in newly abscissed or freshly fallen leaves.
This class of PGR is composed of one chemical compound normally produced in the leaves of plants, originating from chloroplasts, especially when plants are under stress. In general, it acts as an inhibitory chemical compound that affects bud growth, and seed and bud dormancy. It mediates changes within the apical meristem, causing bud dormancy and the alteration of the last set of leaves into protective bud covers. Since it was found in freshly abscissed leaves, it was thought to play a role in the processes of natural leaf drop, but further research has disproven this. In plant species from temperate parts of the world, it plays a role in leaf and seed dormancy by inhibiting growth, but, as it is dissipated from seeds or buds, growth begins. In other plants, as ABA levels decrease, growth then commences as gibberellin levels increase. Without ABA, buds and seeds would start to grow during warm periods in winter and be killed when it froze again. Since ABA dissipates slowly from the tissues and its effects take time to be offset by other plant hormones, there is a delay in physiological pathways that provide some protection from premature growth. It accumulates within seeds during fruit maturation, preventing seed germination within the fruit, or seed germination before winter. Abscisic acid's effects are degraded within plant tissues during cold temperatures or by its removal by water washing in out of the tissues, releasing the seeds and buds from dormancy.[12]
In plants under water stress, ABA plays a role in closing the stomata. Soon after plants are water-stressed and the roots are deficient in water, a signal moves up to the leaves, causing the formation of ABA precursors there, which then move to the roots. The roots then release ABA, which is translocated to the foliage through the vascular system[13] and modulates the potassium and sodium uptake within the guard cells, which then lose turgidity, closing the stomata.[14][15]
ABA exists in all parts of the plant and its concentration within any tissue seems to mediate its effects and function as a hormone; its degradation, or more properly catabolism, within the plant affects metabolic reactions and cellular growth and production of other hormones.[16] Plants start life as a seed with high ABA levels. Just before the seed germinates, ABA levels decrease; during germination and early growth of the seedling, ABA levels decrease even more. As plants begin to produce shoots with fully functional leaves, ABA levels begin to increase, slowing down cellular growth in more "mature" areas of the plant. Stress from water or predation affects ABA production and catabolism rates, mediating another cascade of effects that trigger specific responses from targeted cells. Scientists are still piecing together the complex interactions and effects of this and other phytohormones.
Auxins

The auxin, indole-3-acetic acid
Auxins are compounds that positively influence cell enlargement, bud formation and root initiation. They also promote the production of other hormones and in conjunction with cytokinins, they control the growth of stems, roots, and fruits, and convert stems into flowers.[17] Auxins were the first class of growth regulators discovered.[18] They affect cell elongation by altering cell wall plasticity. They stimulate cambium, a subtype of meristem cells, to divide and in stems cause secondary xylem to differentiate. Auxins act to inhibit the growth of buds lower down the stems (apical dominance), and also to promote lateral and adventitious root development and growth. Leaf abscission is initiated by the growing point of a plant ceasing to produce auxins. Auxins in seeds regulate specific protein synthesis,[19] as they develop within the flower after pollination, causing the flower to develop a fruit to contain the developing seeds. Auxins are toxic to plants in large concentrations; they are most toxic to dicots and less so to monocots. Because of this property, synthetic auxin herbicides including 2,4-D(2,4-dichlorophenoxyacetic) and 2,4,5-T have been developed and used for weed control. Auxins, especially 1-Naphthaleneacetic acid (NAA) and Indole-3-butyric acid (IBA), are also commonly applied to stimulate root growth when taking cuttings of plants. The most common auxin found in plants is indole-3-acetic acid or IAA.
Brassinosteroids

Brassinolide, a major brassinosteroid
Brassinosteroids are a class of polyhydroxysteroids, the only example of steroid based hormones in plants. Brassinosteroids control cell elongation and division, gravitropism, resistance to stress, and xylem differentiation. They inhibit root growth and leaf abscission. Brassinolide was the first identified brassinosteroid and was isolated from extracts of rapeseed (Brassica napus) pollen in 1979.[20]
Cytokinins
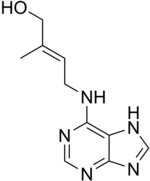
The cytokinin, zeatin
Cytokinins or CKs are a group of chemicals that influence cell division and shoot formation. They were called kinins in the past when the first cytokinins were isolated from yeast cells. They also help delay senescence of tissues, are responsible for mediating auxin transport throughout the plant, and affect internodal length and leaf growth. Cytokinins and auxins often work together, and the ratios of these two groups of plant hormones affect most major growth periods during a plant's lifetime. Cytokinins counter the apical dominance induced by auxins; they in conjunction with ethylene promote abscission of leaves, flower parts, and fruits.[21]
Ethylene

Ethylene
Ethylene is a gas that forms through the breakdown of methionine, which is in all cells. Ethylene has very limited solubility in water and does not accumulate within the cell but diffuses out of the cell and escapes out of the plant. Its effectiveness as a plant hormone is dependent on its rate of production versus its rate of escaping into the atmosphere. Ethylene is produced at a faster rate in rapidly growing and dividing cells, especially in darkness. New growth and newly germinated seedlings produce more ethylene than can escape the plant, which leads to elevated amounts of ethylene, inhibiting leaf expansion (see Hyponastic response). As the new shoot is exposed to light, reactions by phytochrome in the plant's cells produce a signal for ethylene production to decrease, allowing leaf expansion. Ethylene affects cell growth and cell shape; when a growing shoot hits an obstacle while underground, ethylene production greatly increases, preventing cell elongation and causing the stem to swell. The resulting thicker stem can exert more pressure against the object impeding its path to the surface. If the shoot does not reach the surface and the ethylene stimulus becomes prolonged, it affects the stem's natural geotropic response, which is to grow upright, allowing it to grow around an object. Studies seem to indicate that ethylene affects stem diameter and height: When stems of trees are subjected to wind, causing lateral stress, greater ethylene production occurs, resulting in thicker, more sturdy tree trunks and branches. Ethylene affects fruit-ripening: Normally, when the seeds are mature, ethylene production increases and builds-up within the fruit, resulting in a climacteric event just before seed dispersal. The nuclear protein Ethylene Insensitive2 (EIN2) is regulated by ethylene production, and, in turn, regulates other hormones including ABA and stress hormones.[22]
Gibberellins

Gibberellin A1
Gibberellins (GAs) include a large range of chemicals that are produced naturally within plants and by fungi. They were first discovered when Japanese researchers, including Eiichi Kurosawa, noticed a chemical produced by a fungus called Gibberella fujikuroi that produced abnormal growth in rice plants.[23] It was later discovered that GAs are also produced by the plants themselves and they control multiple aspects of development across the life cycle. The synthesis of GA is strongly upregulated in seeds at germination and its presence is required for germination to occur. In seedlings and adults, GAs strongly promote cell elongation. GAs also promote the transition between vegetative and reproductive growth and are also required for pollen function during fertilization.[24]
Jasmonates

Jasmonic acid
Jasmonates (JAs) are lipid-based hormones that were originally isolated from jasmine oil. [25] JAs are especially important in the plant response to attack from herbivores and necrotrophic pathogens.[26] The most active JA in plants is jasmonic acid. Jasmonic acid can be further metabolized into methyl-JA, which is a volatile organic compound. This unusual property means that methyl-JA can act as an airborne signal to communicate herbivore attack to other distant leaves within one plant and even as a signal to neighboring plants. [27] In addition to their role in defense, JAs are also believed to play roles in seed germination, the storage of protein in seeds and root growth. [26]
Salicylic acid

Salicylic acid
Salicylic acid (SA) is a hormone with a related structure to phenol. It was originally isolated from an extract of white willow bark (Salix alba) and is of great interest to human medicine, as it is the precursor of the painkiller, aspirin. In plants, SA plays a critical role in the defense against biotrophic pathogens. In a similar manner to JA, SA can also become methylated. Like methyl-JA, methyl-SA is volatile and can act as a long distance signal to neighboring plants to warn of pathogen attack. In addition to its role in defense, SA is also involved in the response of plants to abiotic stress (particularly drought, temperature, heavy metal and osmotic stress). [28]
Strigolactones

5-deoxystrigol, a strigolactone
Strigolactones (SLs) were originally discovered through studies into the germination of the parasitic weed Striga lutea. It was found that the germination of Striga species was stimulated by the presence of a compound exuded by the roots of its host plant. [29] It was later shown that SLs that are exuded into the soil promote the growth of symbiotic arbuscular mycorrhizal (AM) fungi.[30] More recently, another role of SLs was identified in the inhibition of shoot branching. [31] This discovery of the role of SLs in shoot branching led to a dramatic increase in the interest in these hormones, and it has since been shown that SLs play important roles in leaf senescence, phosphate starvation response, salt tolerance and light signalling. [32]
Other known hormones
Other identified plant growth regulators include:
Plant peptide hormones – encompasses all small secreted peptides that are involved in cell-to-cell signaling. These small peptide hormones play crucial roles in plant growth and development, including defense mechanisms, the control of cell division and expansion, and pollen self-incompatibility.[33] The small peptide CLE25 is known to act as a long-distance signal to communicate water stress sensed in the roots to the stomata in the leaves. [34]
Polyamines – are strongly basic molecules with low molecular weight that have been found in all organisms studied thus far. They are essential for plant growth and development and affect the process of mitosis and meiosis.
Nitric oxide (NO) – serves as signal in hormonal and defense responses (e.g. stomatal closure, root development, germination, nitrogen fixation, cell death, stress response).[35] NO can be produced by a yet undefined NO synthase, a special type of nitrite reductase, nitrate reductase, mitochondrial cytochrome c oxidase or non enzymatic processes and regulate plant cell organelle functions (e.g. ATP synthesis in chloroplasts and mitochondria).[36]
Karrikins – are not plant hormones as they are not produced by plants themselves but are rather found in the smoke of burning plant material. Karrikins can promote seed germination in many species. [37] The finding that plants which lack the receptor of karrikin receptor show several developmental phenotypes (enhanced biomass accumulation and increased sensitivity to drought) have lead some to speculate on the existence of an as yet unidentified karrikin-like endogenous hormone in plants. The cellular karrikin signalling pathway shares many components with the strigolactone signalling pathway.[38]
Triacontanol – a fatty alcohol that acts as a growth stimulant, especially initiating new basal breaks in the rose family. It is found in alfalfa (lucerne), bee's wax, and some waxy leaf cuticles.
Potential medical applications
Plant stress hormones activate cellular responses, including cell death, to diverse stress situations in plants. Researchers have found that some plant stress hormones share the ability to adversely affect human cancer cells. For example, sodium salicylate has been found to suppress proliferation of lymphoblastic leukemia, prostate, breast, and melanoma human cancer cells.[39]Jasmonic acid, a plant stress hormone that belongs to the jasmonate family, induced death in lymphoblastic leukemia cells. Methyl jasmonate has been found to induce cell death in a number of cancer cell lines.
Hormones and plant propagation
Synthetic plant hormones or PGRs are commonly used in a number of different techniques involving plant propagation from cuttings, grafting, micropropagation, and tissue culture.
The propagation of plants by cuttings of fully developed leaves, stems, or roots is performed by gardeners utilizing auxin as a rooting compound applied to the cut surface; the auxins are taken into the plant and promote root initiation. In grafting, auxin promotes callus tissue formation, which joins the surfaces of the graft together. In micropropagation, different PGRs are used to promote multiplication and then rooting of new plantlets. In the tissue-culturing of plant cells, PGRs are used to produce callus growth, multiplication, and rooting.
Seed dormancy
Plant hormones affect seed germination and dormancy by acting on different parts of the seed.
Embryo dormancy is characterized by a high ABA:GA ratio, whereas the seed has a high ABA sensitivity and low GA sensitivity. In order to release the seed from this type of dormancy and initiate seed germination, an alteration in hormone biosynthesis and degradation toward a low ABA/GA ratio, along with a decrease in ABA sensitivity and an increase in GA sensitivity, must occur.
ABA controls embryo dormancy, and GA embryo germination.
Seed coat dormancy involves the mechanical restriction of the seed coat. This, along with a low embryo growth potential, effectively produces seed dormancy. GA releases this dormancy by increasing the embryo growth potential, and/or weakening the seed coat so the radical of the seedling can break through the seed coat.
Different types of seed coats can be made up of living or dead cells, and both types can be influenced by hormones; those composed of living cells are acted upon after seed formation, whereas the seed coats composed of dead cells can be influenced by hormones during the formation of the seed coat. ABA affects testa or seed coat growth characteristics, including thickness, and effects the GA-mediated embryo growth potential. These conditions and effects occur during the formation of the seed, often in response to environmental conditions. Hormones also mediate endosperm dormancy: Endosperm in most seeds is composed of living tissue that can actively respond to hormones generated by the embryo. The endosperm often acts as a barrier to seed germination, playing a part in seed coat dormancy or in the germination process. Living cells respond to and also affect the ABA:GA ratio, and mediate cellular sensitivity; GA thus increases the embryo growth potential and can promote endosperm weakening. GA also affects both ABA-independent and ABA-inhibiting processes within the endosperm.[40]
References
^ https://biology.tutorvista.com/biomolecules/plant-hormones.html
^ http://www.scienceindia.in/home/view_article/70
^ https://www.quora.com/What-are-the-main-functions-of-plant-hormones
^ Went FW, Thimann KV (1937). Phytohormones. New York: The Macmillan Company..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ Tarakhovskaya ER, Maslov Y, Shishova MF (2007). "Phytohormones in algae". Russian Journal of Plant Physiology. 54 (2): 163–170. doi:10.1134/s1021443707020021.
^ "Gibberellin formation in microorganisms". Plant Growth Regulation. 15: 303–314. doi:10.1007/BF00029903.
^ Srivastava LM (2002). Plant growth and development: hormones and environment. Academic Press. p. 140. ISBN 0-12-660570-X.
^ Öpik H, Rolfe SA, Willis AJ, Street HE (2005). The physiology of flowering plants (4th ed.). Cambridge University Press. p. 191. ISBN 978-0-521-66251-2.
^ Swarup R, Perry P, Hagenbeek D, Van Der Straeten D, Beemster GT, Sandberg G, Bhalerao R, Ljung K, Bennett MJ (July 2007). "Ethylene upregulates auxin biosynthesis in Arabidopsis seedlings to enhance inhibition of root cell elongation". The Plant Cell. 19 (7): 2186–96. doi:10.1105/tpc.107.052100. PMC 1955695. PMID 17630275.
^ Srivastava 2002, p. 143
^ Thomas Elliot W, Thomas L R (1979). Botany: a brief introduction to plant biology. New York: Wiley. pp. 155–170. ISBN 0-471-02114-8.
^ Feurtado JA, Ambrose SJ, Cutler AJ, Ross AR, Abrams SR, Kermode AR (February 2004). "Dormancy termination of western white pine (Pinus monticola Dougl. Ex D. Don) seeds is associated with changes in abscisic acid metabolism". Planta. 218 (4): 630–9. doi:10.1007/s00425-003-1139-8. PMID 14663585.
^ Ren H, Gao Z, Chen L, Wei K, Liu J, Fan Y, Davies WJ, Jia W, Zhang J (2007). "Dynamic analysis of ABA accumulation in relation to the rate of ABA catabolism in maize tissues under water deficit". Journal of Experimental Botany. 58 (2): 211–9. doi:10.1093/jxb/erl117. PMID 16982652. Archived from the original on 2012-07-10.
^ Else MA, Coupland D, Dutton L, Jackson MB (January 2001). "Decreased root hydraulic conductivity reduces leaf water potential, initiates stomatal closure, and slows leaf expansion in flooded plants of castor oil (Ricinus communis) despite diminished delivery of ABA from the roots to shoots in xylem sap". Physiologia Plantarum. 111 (1): 46–54. doi:10.1034/j.1399-3054.2001.1110107.x.
^ Yan J, Tsuichihara N, Etoh T, Iwai S (October 2007). "Reactive oxygen species and nitric oxide are involved in ABA inhibition of stomatal opening". Plant, Cell & Environment. 30 (10): 1320–5. doi:10.1111/j.1365-3040.2007.01711.x. PMID 17727421.
^
Kermode AR (December 2005). "Role of Abscisic Acid in Seed Dormancy". J Plant Growth Regul. 24 (4): 319–344. doi:10.1007/s00344-005-0110-2.
^ Osborne DJ, McManus MT (2005). Hormones, signals and target cells in plant development. Cambridge University Press. p. 158. ISBN 978-0-521-33076-3.
^ Classification of auxin related compounds based on similarity of their interaction fields: Extension to a new set of compounds Tomic, S.1,2, Gabdoulline, R.R.1, Kojic-Prodic, B.2 and Wade, R.C.11 European Molecular Biology Laboratory, 69012 Heidelberg, Germany
2Institute Rudjer Boskovic, HR-10000 Zagreb, Croatia
^ Walz A, Park S, Slovin JP, Ludwig-Müller J, Momonoki YS, Cohen JD (February 2002). "A gene encoding a protein modified by the phytohormone indoleacetic acid". Proceedings of the National Academy of Sciences of the United States of America. 99 (3): 1718–23. Bibcode:2002PNAS...99.1718W. doi:10.1073/pnas.032450399. PMC 122257. PMID 11830675.
^ Grove MD, Spencer GF, Rohwedder WK, Mandava N, Worley JF, Warthen JD, et al. (1979). "Brassinolide, a plant growth-promoting steroid isolated from Brassica napus pollen". Nature. 281 (5728): 216–217. Bibcode:1979Natur.281..216G. doi:10.1038/281216a0.
^
Sipes DL, Einset JW (August 1983). "Cytokinin stimulation of abscission in lemon pistil explants". J Plant Growth Regul. 2 (1–3): 73–80. doi:10.1007/BF02042235.
^ Wang Y, Liu C, Li K, Sun F, Hu H, Li X, et al. (August 2007). "Arabidopsis EIN2 modulates stress response through abscisic acid response pathway". Plant Molecular Biology. 64 (6): 633–44. doi:10.1007/s11103-007-9182-7. PMID 17533512.
^ Grennan AK (June 2006). "Gibberellin metabolism enzymes in rice". Plant Physiology. 141 (2): 524–6. doi:10.1104/pp.104.900192. PMC 1475483. PMID 16760495.
^ Tsai FY, Lin CC, Kao CH (January 1997). "A comparative study of the effects of abscisic acid and methyl jasmonate on seedling growth of rice". Plant Growth Regulation. 21 (1): 37–42. doi:10.1023/A:1005761804191.
^ Demole E, Lederer E, Mercier D (1962). "Isolement et détermination de la structure du jasmonate de méthyle, constituant odorant caractéristique de l'essence de jasmin" [Isolation and determination of the structure of methyl jasmonate, a fragrant constituent characteristic of jasmine oil]. Helvetica Chimica Acta (in French). 45 (2): 675–685. doi:10.1002/hlca.19620450233.
^ ab Browse J (2005). "Jasmonate: an oxylipin signal with many roles in plants". Vitamins and Hormones. 72: 431–56. doi:10.1016/S0083-6729(05)72012-4. PMID 16492478.
^ Katsir L, Chung HS, Koo AJ, Howe GA (August 2008). "Jasmonate signaling: a conserved mechanism of hormone sensing". Current Opinion in Plant Biology. 11 (4): 428–35. doi:10.1016/j.pbi.2008.05.004. PMC 2560989. PMID 18583180.
^ Rivas-San Vicente M, Plasencia J (June 2011). "Salicylic acid beyond defence: its role in plant growth and development". Journal of Experimental Botany. 62 (10): 3321–38. doi:10.1093/jxb/err031. PMID 21357767.
^ Xie X, Yoneyama K, Yoneyama K (2010-07-01). "The strigolactone story". Annual Review of Phytopathology. 48 (1): 93–117. doi:10.1146/annurev-phyto-073009-114453. PMID 20687831.
^ Akiyama K, Matsuzaki K, Hayashi H (June 2005). "Plant sesquiterpenes induce hyphal branching in arbuscular mycorrhizal fungi". Nature. 435 (7043): 824–7. Bibcode:2005Natur.435..824A. doi:10.1038/nature03608. PMID 15944706.
^ Gomez-Roldan V, Fermas S, Brewer PB, Puech-Pagès V, Dun EA, Pillot JP, Letisse F, Matusova R, Danoun S, Portais JC, Bouwmeester H, Bécard G, Beveridge CA, Rameau C, Rochange SF (September 2008). "Strigolactone inhibition of shoot branching". Nature. 455 (7210): 189–94. Bibcode:2008Natur.455..189G. doi:10.1038/nature07271. PMID 18690209.
^ Saeed W, Naseem S, Ali Z (2017-08-28). "Strigolactones Biosynthesis and Their Role in Abiotic Stress Resilience in Plants: A Critical Review". Frontiers in Plant Science. 8: 1487. doi:10.3389/fpls.2017.01487. PMC 5581504. PMID 28894457.
^ Lindsey K, Casson S, Chilley P (February 2002). "Peptides: new signalling molecules in plants". Trends in Plant Science. 7 (2): 78–83. doi:10.1016/S0960-9822(01)00435-3. PMID 11832279.
^ Takahashi F, Suzuki T, Osakabe Y, Betsuyaku S, Kondo Y, Dohmae N, Fukuda H, Yamaguchi-Shinozaki K, Shinozaki K (April 2018). "A small peptide modulates stomatal control via abscisic acid in long-distance signalling". Nature. 556 (7700): 235–238. Bibcode:2018Natur.556..235T. doi:10.1038/s41586-018-0009-2. PMID 29618812.
^ Shapiro AD (2005). "Nitric oxide signaling in plants". Vitamins and Hormones. 72: 339–98. doi:10.1016/S0083-6729(05)72010-0. PMID 16492476.
^ Roszer T (2012). "Nitric Oxide Synthesis in the Chloroplast". In Roszer T. The Biology of Subcellular Nitric Oxide. New York, London, Heidelberg: Springer. ISBN 978-94-007-2818-9.
^ Chiwocha SD, Dixon KW, Flematti GR, Ghisalberti EL, Merritt DJ, Nelson DC, et al. (2009-10-01). "Karrikins: A new family of plant growth regulators in smoke". Plant Science. 177 (4): 252–256. doi:10.1016/j.plantsci.2009.06.007.
^ Li W, Nguyen KH, Chu HD, Ha CV, Watanabe Y, Osakabe Y, Leyva-González MA, Sato M, Toyooka K, Voges L, Tanaka M, Mostofa MG, Seki M, Seo M, Yamaguchi S, Nelson DC, Tian C, Herrera-Estrella L, Tran LP (November 2017). "The karrikin receptor KAI2 promotes drought resistance in Arabidopsis thaliana". PLoS Genetics. 13 (11): e1007076. doi:10.1371/journal.pgen.1007076. PMC 5703579. PMID 29131815.
^ Fingrut O, Flescher E (April 2002). "Plant stress hormones suppress the proliferation and induce apoptosis in human cancer cells". Leukemia. 16 (4): 608–16. doi:10.1038/sj.leu.2402419. PMID 11960340.
^ Gerhard W (2000). "Seed Dormancy". The Seed Biology Place. Royal Holloway University of London.
External links
| Wikimedia Commons has media related to Plant hormones. |
Simple plant hormone table with location of synthesis and effects of application — this is the format used in the descriptions at the ends of the Wikipedia articles on individual plant hormones.
Hormonal Regulation of Gene Expression and Development — Detailed intro including genetic information.
Botany | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
History of botany | |||||||||||
| Subdisciplines |
| ||||||||||
Plant groups |
| ||||||||||
Plant morphology .mw-parser-output .nobold{font-weight:normal} (glossary) |
| ||||||||||
|
| ||||||||||
| Plant growth and habit |
| ||||||||||
Reproduction
|
| ||||||||||
| Plant taxonomy |
| ||||||||||
| Practice |
| ||||||||||
|
| ||||||||||
| |||||||||||
Authority control |
|
|---|


